L’épigénétique, contributeur d’expression
Etienne Bucher

L’épigénétique est un domaine de recherche récent qui excite de nombreux scientifiques à cause de son potentiel dans l’amélioration des plantes et la création variétale. Le but de ce texte est d’expliquer ce qu’est l’épigénétique, placer cette réalité dans un contexte historique, évaluer sa valeur et ses limites pour l’amélioration des plantes et évoquer les implications de ce concept sur l’évolution des plantes et d’autres organismes.
La génétique, et les règles de l’hérédité des caractères qui en découlent, ont été décrites la première fois par Gregor Mendel (1822-1884), un moine autrichien. La signification de ses découvertes ne fut appréciée que trois décennies plus tard et établissait la logique et la possibilité de pouvoir prédire la transmission de caractères d’une génération à l’autre. On appelle cela l’hérédité mendélienne. Puisque la génétique permet de prédire le résultat d’hybridations, elle est un outil indispensable dans l’obtention de nouvelles variétés. Par contre, la génétique a ses limites : elle n’explique pas toute la variabilité des caractères observée. En effet, même si le génome humain a été complètement décodé, c’est toujours un défi majeur d’identifier les gènes qui influencent des caractères importants, un problème intitulé l’hérédité manquante (Gribbin, 2008).
L’humain plus complexe que l’arabette
Suite au séquençage des génomes complets, il est démontré qu’il n’existe pas de corrélation entre la complexité d’un organisme, la taille de son génome et le nombre de gènes qu’il contient. Par exemple, le génome humain contient moins de 20 000 gènes alors que la petite plante modèle nommée arabette (Arabidopsis thaliana) en contient plus de 27 000 (Figure 1) et cela, alors que le génome de l’arabette est 22 fois plus petit que le génome humain ! Cela veut dire que les génomes peuvent contenir plus ou moins de « matière noire » dont on ne connaît pas encore la fonction. Ces dernières années, des chercheurs ont proposé que la matière noire du génome soit un contributeur majeur. Si on assume que l’humain est plus complexe que l’arabette, il faut en conclure qu’il existe un facteur autre que le nombre de gènes qui rend un organisme plus complexe. La régulation de l’expression des gènes doit y contribuer. Et c’est là que l’épigénétique entre en jeu.
Le code épigénétique dorénavant lisible
L’épigénétique se définit par la transmission de l’état d’expression d’un gène (ON ou OFF, Figure 2) d’une génération à l’autre, indépendamment des différences génétiques. Cela signifie que deux organismes contenant exactement le même génome, par exemple de vrais jumeaux ou des clones, peuvent présenter des caractères différents parce qu’ils expriment différemment les mêmes gènes. Ceci est très important car implique que le phénotype d’un organisme n’est pas seulement le résultat du code génétique mais aussi du code épigénétique. Les modifications épigénétiques sont matérialisées par des marques biochimiques ajoutées aux protéines associées à l’ADN ou à la molécule d’ADN elle-même. Dépendant de leur nature, ces marques peuvent réprimer ou activer les gènes. Grâce aux nouvelles technologies de séquençage, il est maintenant possible de lire le code épigénétique entier d’une plante. Cela aide les chercheurs à mieux comprendre comment ces marques influencent l’expression des gènes et par cela les caractères d’une plante.
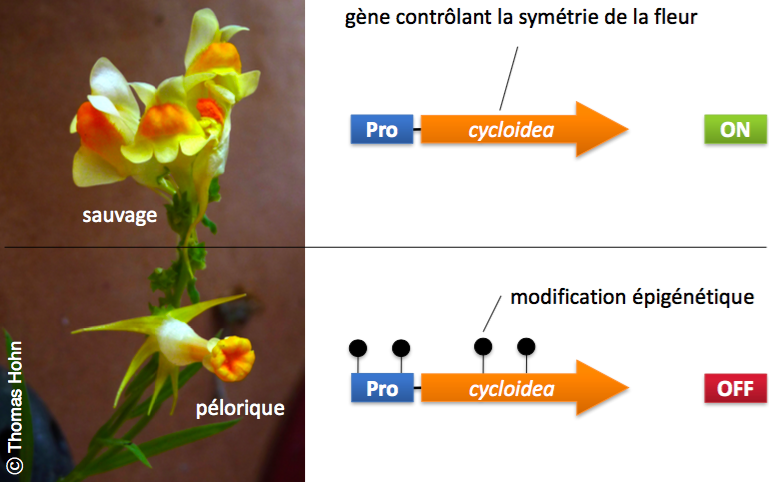
(Linaria vulgaris). En haut: la fleur sauvage (ou prédominante), le gène cycloidea est actif, ce qui permet un développement normal de la fleur. En bas: la fleur pélorique, le gène cycloidea est inactivé par la présence de marques épigénétiques changeant la forme de la fleur.
Effets maternels et paternels
Deux critères différencient fortement la génétique de l’épigénétique : (i) la mutation génétique est quasi irréversible, alors que les modifications épigénétiques sont réversibles et (ii) il y a des indications que le code épigénétique peut être influencé par l’environnement, alors que les mutations génétiques sont rares et requièrent un agent mutagène, par exemple les radiations naturelles de type UVs ou les radiations artificielles de type rayons X ou gamma. Que l’environnement puisse potentiellement influencer le code épigénétique d’une plante a de fortes implications : cela voudrait dire qu’une plante peut se souvenir d’un stress qu’elle a subi et transférer cette mémoire à la prochaine génération. Intuitivement, cela paraît logique car, en utilisant cette possibilité de transfert d’information, la prochaine génération pourra être mieux préparée aux stress que ses parents ont dû subir. De nombreuses études ont démontré des effets maternels et paternels quand les plantes ont subi des stress environnementaux comme la lumière, la température ou la qualité du sol (Youngson and Whitelaw 2008). Bien que ces effets soient intéressants, ils ne sont pas stables d’un point de vue de l’héritabilité. Dans quasiment tous les cas décrits dans la littérature, l’effet est perdu après deux générations. Ces effets maternels et paternels ne constituent pas une mémoire épigénétique dans le sens strict.
Un rôle important dans l’expression de caractères
Et pourtant il existe de très beaux exemples chez les plantes, où une mémoire épigénétique peut être transmise de manière stable et cela même pendant des décennies. La linaire commune (Linaria vulgaris) peut exister sous deux formes produisant des fleurs à symétrie bilatérale (type prédominant) et des fleurs à symétrie radiale (péloriques, Figure 2). Carl von Linné avait déjà mentionné cette observation en 1749 et indiqué son étonnement que deux plantes identiques puissent produire deux types de fleurs. C’est pour cela qu’il a appelé la fleur de symétrie radiale « pélorique » (ce qui veut dire « monstrueux » en grec). Aujourd’hui on sait que les deux types de fleurs diffèrent simplement dans l’état épigénétique du gène cycloidea (Cubas et al. 1999). Depuis, plusieurs exemples où le code épigénétique d’un gène spécifique peut influencer des caractères d’importance agronomique ont été décrits : les variantes somaclonales chez le palmier à huile qui affectent le développement du fruit (Ong-Abdullah et al. 2015) et la définition du sexe des fleurs chez le melon (https://www.jardinsdefrance.org/concombre-melon-la-sexualite-devoilee/). Bien que ces exemples démontrent clairement que l’épigénétique peut jouer un rôle très important dans l’expression de caractères, il n’est pour l’instant pas encore possible d’évaluer l’étendue effective de cette régulation. Est-ce que se sont seulement des exceptions, ou est-ce que le contrôle épigénétique de caractères différents est la règle chez les plantes ?
Des questions subsistent
Finalement, les questions essentielles sont les suivantes: est-il possible de modifier l’épigénome d’une plante de manière héritable pour que ce soit utile pour l’amélioration des plantes ? Est-ce qu’il existe des stress ou autres conditions qui peuvent induire de la variabilité épigénétique stable ? Ce sont aussi des questions fondamentales importantes car leur réponse nous permettra de conclure si l’évolution est un phénomène purement aléatoire suivi par la sélection ou s’il existe un moyen pour les plantes – à travers l’épigénétique – d’évoluer d’une manière dirigée. On se trouve donc au début de cette recherche fascinante et prometteuse.
A lire …
– Cubas P, Vincent C, Coen E. 1999. An epigenetic mutation responsible for natural variation in floral symmetry. Nature 401: 157–161.
– Gribbin S. 2008. The case of the missing heritability. ?1–4.
Ong-Abdullah M, Ordway JM, Jiang N, Ooi S-E, Kok S-Y, Sarpan N, Azimi N,
– Hashim AT, Ishak Z, Rosli SK, et al. 2015. Loss of Karma transposon methylation underlies the mantled somaclonal variant of oil palm. Nature 525: 533–537.
– Youngson NA, Whitelaw E. 2008. Transgenerational epigenetic effects. Annu Rev Genom Hum G 9: 233–25