Inflorescences des Berberis, une diversité de formes
Alain Cadic
Les inflorescences des berbéridacées ligneuses ( Berberis, Mahonia, X Mahoberberis et Nandina) se rangent dans la catégorie des inflorescences racémeuses. Chez les Berberis ce type se décline en de nombreuses formes liées à l’évolution du genre au cours de sa dispersion dans de nombreuses régions du monde.
Le genre Berberis est apparu très anciennement, des fossiles datant du Paléocène (- 66 à – 56 millions d’années) en ont été découverts dans le nord-est de la Chine. Son expansion ultérieure sur l’ensemble de l’Eurasie et de l’Amérique s’est accompagnée d’une grande diversification des caractéristiques morphologiques des 500 espèces ou plus qui le constituent, parmi lesquelles la forme de l’inflorescence, le nombre de fleurs et de fruits. Ce nombre varie de 1 à 250 fleurs (photos 1 et 2).


Le modèle général : la grappe
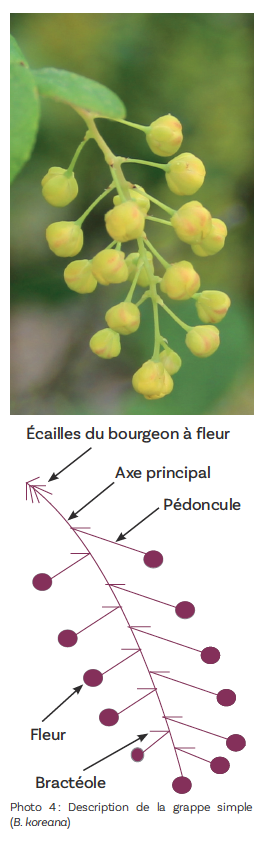
La multiplicité des types inflorescentiels se retrouve dans le vocabulaire employé pour tenter de caractériser chacune des espèces : fleur solitaire, inflorescence fasciculée, en ombelle, en grappe, inflorescence pseudo-ombellée, en épi, en panicule.
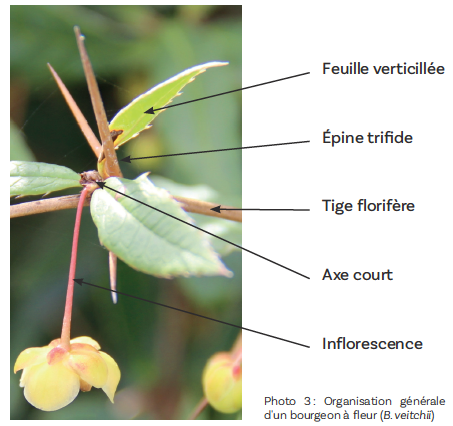
Chacune des inflorescences est entourée de feuilles caduques ou persistantes portées par un axe court lui-même axillé par une feuille transformée en « épine » plus ou moins développée (photo 3).
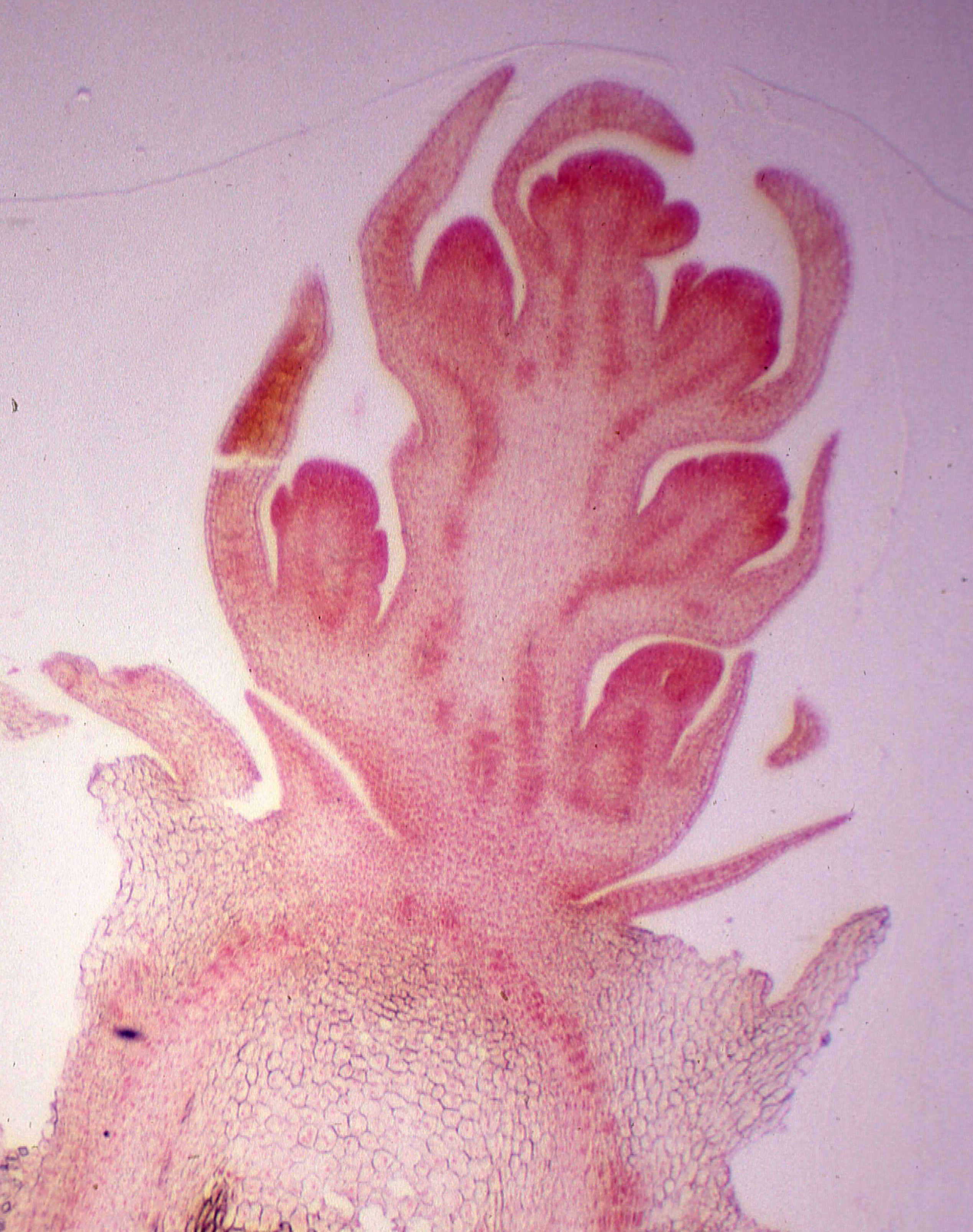
Le modèle général d’inflorescence du Berberis est la grappe dont les éléments essentiels peuvent se réduire ou encore, à l’opposé, se multiplier (photo 4). Cette structure s’observe nettement sur des coupes longitudinales de bourgeons à fleurs observées en microscopie (photo 5).
fleur de B. darwinii – © Inra – A. Cadic et D. Martin
Une structure dépendant de deux mécanismes
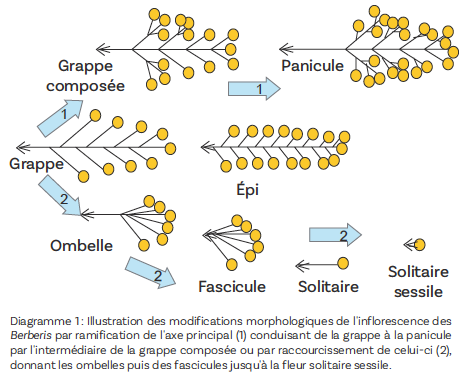
Deux mécanismes principaux contribuent à la modification de la structure de l’inflorescence (diagramme 1).
Le raccourcissement général de l’axe principal s’accompagne de la diminution du nombre de fleurs. Celui de la partie apicale de l’axe conduit à une forme d’ombelle ou de pseudo-ombelle comme chez B. amoena ou B. sieboldii. L’absence d’axe central sans diminution de longueur des pédicelles floraux donne des inflorescences de type fasciculé comme chez B. juliane (photo 6). La réduction du nombre de fleurs conduit à la production de structures uniflores comme chez B. veitchii (photo 7) qui devient une fleur sessile si, de plus, le pédicelle floral disparaît comme chez B. dictyophylla (photo 1).
À l’opposé, l’allongement de l’axe principal s’accompagne de l’augmentation du nombre de fleurs. Il peut s’accompagner d’un début de ramification basale donnant d’abord des grappes composées (B. dielsiana) puis de véritables panicules (B. paniculata) constituées de grappes de grappes ou de grappes d’ombelles (B. aggregata).

B. veitchii – © Inra – A. Cadic

Un genre exceptionnel pour ses inflorescences
Compte tenu de la conservation des gènes, il pourrait être intéressant d’analyser les analogues des gènes déjà séquencés sur les espèces modèles, d’analyser et comparer les mutations qui les affectent en prenant en compte des modèles d’inflorescences contrastés qui caractérisent le genre Berberis, un genre assez exceptionnel de ce point de vue.
* Autogamie : reproduction par autofécondation préférentielle ou stricte favorisée, chez les Berberis, par le processus de thigmonastie provoquant le rabat vif des clapets porteurs de pollen sur la couronne stigmatique réceptrice lorsque les insectes viennent prélever le nectar sécrété à la base des anthères.