Arômes et parfums, le dilemme des plantes à fleurs
Jean-Claude Caissard
Il existe de nombreux points communs entre les mécanismes de défenses face aux insectes phytophages et les mécanismes d’attraction des insectes pollinisateurs. Qu’il s’agisse de repousser ou d’attirer, la plante émet des composés organiques volatils ou COV[1]. Il s’agit de molécules passant spontanément à l’état gazeux sous pression atmosphérique et température ambiante. Ces molécules sont produites par tous les êtres vivants et elles sont principalement impliquées dans la communication entre les organismes.
Des composés attractifs et répulsifs
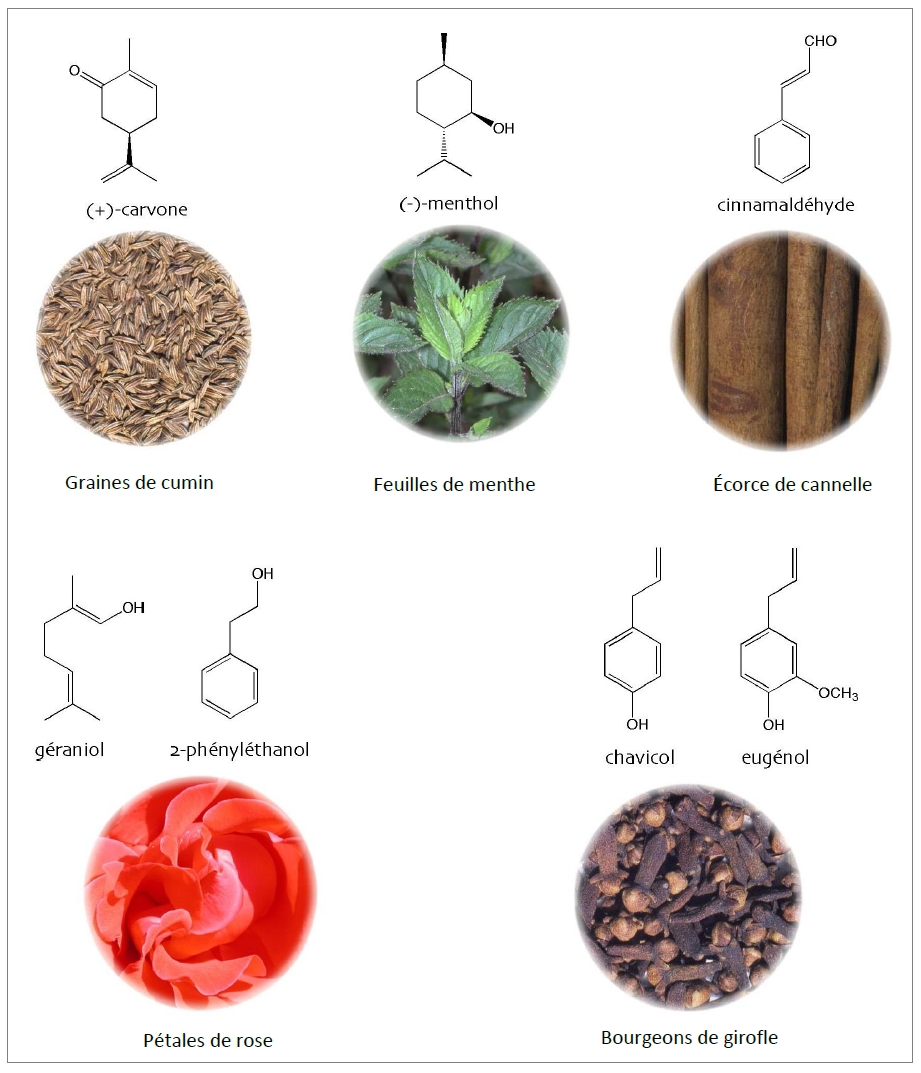
Chez les végétaux, la biosynthèse des COV est souvent assurée par des cellules spécialisées dans la sécrétion. Ces cellules sont situées soit au niveau des fleurs, et on parle alors de plante à parfum, soit au niveau de l’appareil végétatif, et on parle alors de plante aromatique. Parmi les COV, on peut citer par exemple (Fig. 1) le menthol, stocké dans les glandes de menthe poivrée, Mentha x piperita – Lamiaceae -, l’eugénol et le chavicol émis par le bouton floral de girofle, Eugenia aromatica – Myrtaceae –, le géraniol et le 2-phényléthanol sécrétés par les épidermes des pétales de rose, Rosa x hybrida – Rosaceae –, le cinnamaldéhyde synthétisé par l’écorce interne du tronc de cannelle, Cinnamomum verum – Lauraceae –, ou encore la carvone produite par le tégument des diakènes du cumin des prés, Carum carvi – Apiaceae. La plupart sont des produits du métabolisme secondaire, c’est-à-dire qu’ils ne participent pas aux fonctions essentielles (respiration, photosynthèse…) comme les métabolites primaires. Depuis quelques années, ces métabolites sont dits « spécialisés » pour les opposer aux métabolites primaires « généraux » et surtout pour marquer leur rôle dans les interactions avec les autres organismes.
Les COV peuvent être toxiques pour les bactéries, les champignons, et apéritifs, addictifs ou anorexiques pour les insectes et les vertébrés. Les deux effets majeurs sont cependant l’attraction des pollinisateurs et la répulsion des phytophages et parasites.
Une reconnaissance complexe
La reconnaissance d’une odeur est complexe, elle tient autant de l’inné que de l’acquis. Il est, par exemple, possible d’apprendre à un cricket que l’odeur du menthol, habituellement répulsive, est associée à de la nourriture. A l’opposé, chez certaines orchidées, les fleurs émettent des phéromones d’insecte femelle mais aucune récompense pour le mâle, ni nourriture ni lieu de reproduction. Ces fleurs sont dites décevantes ou trompeuses pour l’insecte, qui continue pourtant à être immanquablement attiré à un point tel que Darwin écrivit « it appears to me quite incredible that the same insect should go on visiting flower after flower of these Orchids, although it never obtains any nectar »[2]
Chez le papillon (ex : Lymantria dispar –Lymantriidae) les antennes sont couvertes d’organes sensoriels pourvus de pores. Les COV passent par ces pores et sont captés par de petites protéines. Le complexe est transporté jusqu’à des récepteurs membranaires, la transduction du signal provoque alors un stimulus interprété par le cerveau.
L’homme aux 10 000 odeurs
Dans le nez humain, l’air inspiré passe sur la muqueuse nasale et plus particulièrement sur l’épithélium olfactif. L’explication complète du mécanisme de l’olfaction a été réalisée par les prix Nobel de 2004, R. Axel et L. Buck (nobelprize.org). Les quelques 10 millions de neurones olfactifs de cet épithélium se terminent par des cils sensoriels porteurs de protéines chimio réceptrices. Le cerveau associe alors une odeur à chaque combinaison d’informations et peut ainsi interpréter près de 10 000 odeurs. Mais les seuils de détection diffèrent selon les VOC. Il est par exemple possible de détecter une odeur proche de celle de la thiamine avec 0,000002 µg de bis (2-méthyl-3-furyl) disulfide dans 1 litre d’eau alors qu’il faut monter à 75 µg/l pour sentir l’odeur de rose du géraniol.
Molécules et sécrétion
L’extraordinaire diversification des COV semble pourtant ne trouver sa source que dans quelques familles de molécules dont les dérivés terpéniques, les dérivés aromatiques et les dérivés d’acides gras. La biosynthèse de ces COV commence à être bien connue mais leur trafic intracellulaire reste une énigme et leur excrétion cellulaire rarement documentée. Par exemple, dans les trichomes glandulaires d’Artemisia annua, les vésicules du reticulum endoplasmique[3] fusionnent avec la membrane plasmique et se déversent à l’extérieur de la cellule dans un mécanisme dit d’exocytose. Le transport des COV peut aussi s’effectuer directement à travers la membrane. C’est le cas par exemple chez Nepeta racemosa, chez Mentha x piperita et même chez Leonitis leonurus où des vésicules ont pourtant été détectées mais sans aucune figure de fusion avec la membrane plasmique.
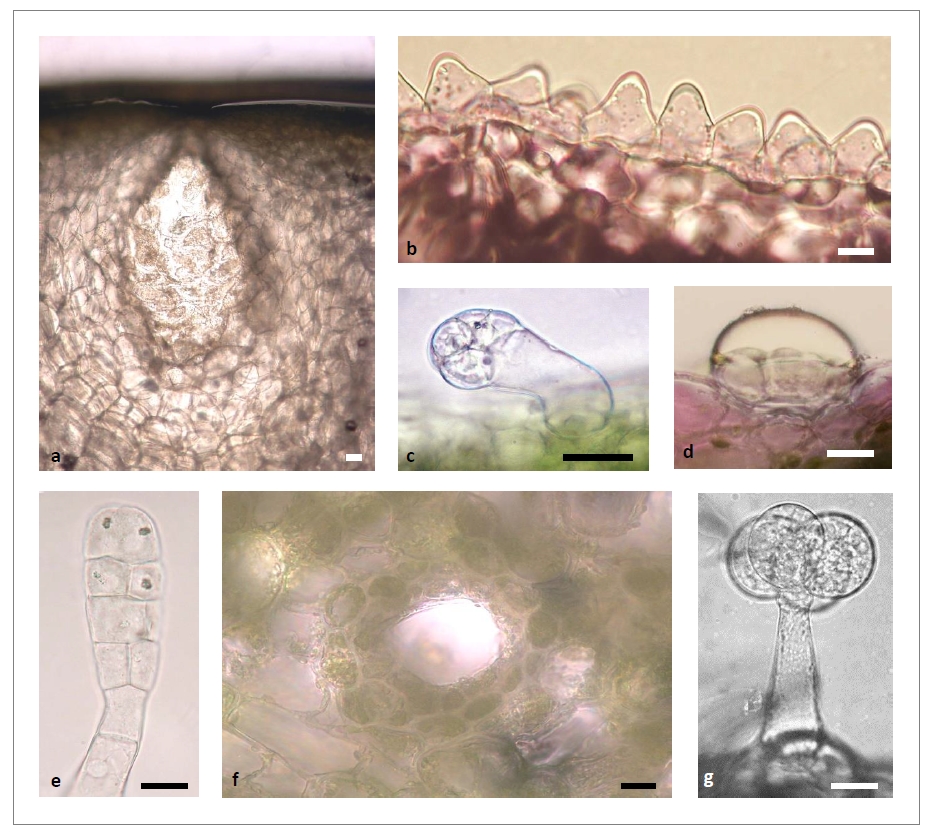
a, poche sécrétrices du péricarpe du fruit de Citrus sp. ; b, cellules papilleuses de l’épiderme de pétale de Lamium purpureum ; c, hydathode de feuille de Nicotiana sylvestris ; d, glande peltée de feuille de Mentha x piperita ; e, trichomes sécréteur à tête pluricellulaire et cristaux d’oxalate de calcium de pétale de N. sylvestris ; f, canal résinifère de branche de Pseudotsuga menziesii ; g, trichome sécréteur à 4 cellules de tête de feuille de Lycopersicon esculentum. Barres d’échelle, 10 µm sauf a, d et f, 20 µm.
Des cellules sécrétrices spécifiques

a, micrographie électronique environnementale de l’épiderme supérieur montrant des gouttelettes de sécrétion ; b, vue paradermale des cellules coniques de l’épiderme supérieur
Toutes les cellules semblent capables d’excrétés des COV en plus ou moins grande quantité. Néanmoins, l’usage est de qualifier de « cellules sécrétrices » toute cellule à phénotype particulier, excrétant ou accumulant des composés toxiques et/ou des composés du métabolisme secondaire (Fig. 2). On trouve ainsi des nectaires sécrétant principalement des polysaccharides, des hydathodes sécrétant de l’eau et des composés hydrosolubles, des cellules papilleuses sécrétant des COV, des canaux et des poches sécrétant des terpènes dans des espaces intercellulaires, des trichomes glandulaires sécrétant des métabolites secondaires ou des sucs digestifs etc. Les cellules papilleuses des pétales ont une forme conique. Ce type de cellules compose très souvent l’épiderme supérieur des pétales. C’est le cas par exemple chez Rosa x hybrida (Fig. 3), chez Erysimum cheiri, ou chez Petunia x hybrida. Parfois, les cellules coniques sont regroupées en « osmophores » comme chez beaucoup d’Orchidées chez lesquelles elles semblent guider les insectes pollinisateurs dans la fleur.
Langage écologique
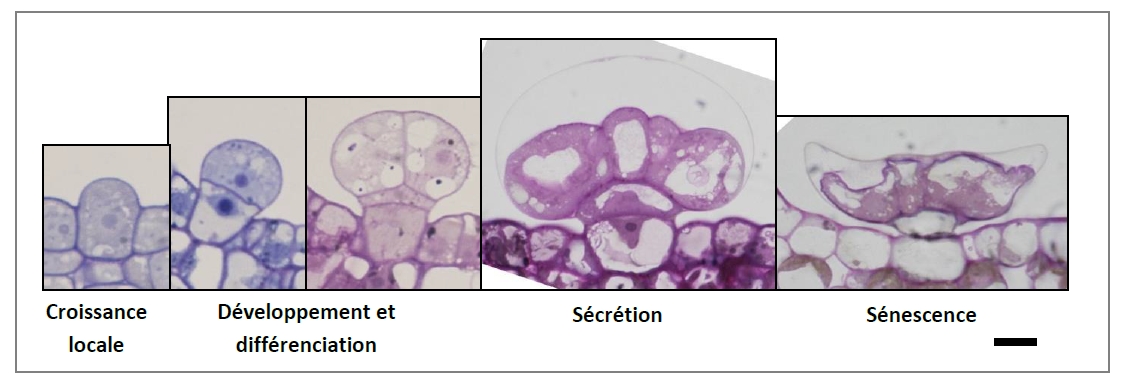
Très souvent, les trichomes sécréteurs, les poches et les canaux sont trouvés dans les parties vertes à protéger (feuilles, tiges, sépales…). C’est le cas par exemple des Lamiaceae avec leurs glandes capitées et leurs glandes peltées. Ces dernières, gonflées par des terpènes dans un espace extra-cellulaire sous-cuticulaire, éclatent au contact d’une patte d’insecte.
Les cellules sécrétrices sont donc les vecteurs des messages portés par les COV qui constituent eux-mêmes un véritable langage écologique. Ce monde microscopique et chimique révèlera encore certainement bien des surprises.
[1] COV pour Composés Organiques Volatils
[2] « Il me semble assez incroyable que le même insecte puisse visiter ces Orchidées, fleurs après fleurs, alors qu’il n’y obtiendra aucun nectar ».
[3] Le reticulum endoplasmique est un organite cellulaire capable, entre autre, de transporter certaines substances via des vésicules.