Domestication et diversification des plantes : le cas de la vigne
La vigne, Vitis vinifera L., a été initialement domestiquée à partir de son ancêtre sauvage, V. vinifera subsp. sylvestris dans une zone allant du sud du Caucase à la Mésopotamie. Des changements morphologiques et adaptatifs ont eu lieu lors de cette étape (syndrome de domestication), mais également tout au long du lent processus de sélection, réalisé par l’homme, tout d’abord de façon spontanée, puis par sélection dirigée, à partir de la fin du XVIIIe siècle.

La vigne a été modifiée pour des caractères morphologiques majeurs (architecture et port des rameaux, fleurs, baies, grappes) et d’adaptation à l’environnement (vigueur et phénologie notamment), mais peu pour les stress biotiques. Vitis vinifera L. est la principale espèce du genre Vitis utilisée pour la production de raisins, qui sont vinifiés, consommés en frais ou transformés en raisins secs. Le principal produit de la vigne reste cependant le vin et ses alcools dérivés, issus de la fermentation des raisins sous l’action des levures du genre Saccharomyces. Des variétés issues, par hybridation avec d’autres espèces du genre Vitis, sont également cultivées pour la production de raisin ou de vin (par exemple la variété Isabelle hybride entre V. vinifera et V. labrusca). Les espèces de Vitis (V. riparia, V. rupestris et V. berlandieri) ont été aussi utilisées pour la création de porte-greffes, après l’introduction du phylloxera à la fin du XIXe siècle en Europe.
Vitis vinifera et son ancêtre sauvage : une domestication ancienne
L’espèce V. vinifera L., euro-asiatique, a été domestiquée à partir de V. vinifera subsp. sylvestris (Gmelin) Hegi, encore appelée lambrusque. La domestication a eu lieu dans une zone allant du sud du Caucase à la Mésopotamie où ont été découverts les premiers indices de mise en culture (notamment des restes d’acide tartrique dans des amphores), datant du VIe au Ve millénaire avant J.-C. À partir de cette zone de domestication primaire, la culture de la vigne a été propagée vers l’ouest, sur le pourtour de la Méditerranée, en accompagnant les principales civilisations (Égyptiens, Phéniciens, Grecs, Romains…), et à l’est vers l’Inde et la Chine. Des évènements de domestication secondaire ont également été identifiés dans les zones où la vigne a été introduite. Elle a ensuite progressé vers le nord de l’Europe et les pays du Nouveau Monde. Elle a évolué grâce à la multiplication sexuée, mais le maintien des variétés une fois créées a été rendu possible par la multiplication végétative.
Les principales modifications liées à la domestication
La domestication de la vigne par l’homme a eu comme principal objectif de garantir une production régulière de raisins de qualité faciles à récolter. À l’état sauvage, la vigne est une liane qui grimpe dans les arbres, ce qui rend difficile la cueillette des raisins. Les grains sont très petits, très acides, peu juteux et de couleur noire. La vigne sauvage est dioïque (1*). Seuls les pieds femelles à proximité de pieds mâles peuvent donc produire des raisins. Les principaux effets de la domestication ont ainsi modifié l’architecture des rameaux, la fleur, et les fruits.
Concernant l’architecture, d’une liane grimpant dans les arbres, la culture de la vigne a évolué vers le mode de conduite que l’on connaît actuellement en passant probablement par des intermédiaires, telle la conduite appelée « arbustum » (2*). Les modes de conduite actuels sont très variés, comme le gobelet dans les vignobles méditerranéens, la pergola pour la production de raisin de table ou encore certaines tailles très spécifiques comme le « kouloura » de l’île de Santorin ou le mode de culture de Lanzarote (Canaries), permettent une adaptation de la vigne à des vents extrêmes. Au niveau morphologique, des différences existent sur l’ensemble des organes de la plante (feuille, fleur, pépin, grappe, baie) (Figure n° 1).
Parmi les plus importantes, il faut signaler l’apparition de l’hermaphrodisme qui permet notamment la pollinisation de la fleur par son propre pollen et donc une régularité de production. De même, la baie et la grappe sont deux organes ayant subi un syndrome de domestication très marqué : la domestication puis la sélection ont conduit à des variétés de vigne présentant des baies plus grosses, plus sucrées et plus juteuses (Figure n° 2). La taille des grappes a également augmenté. La nouaison plus régulière, permet un développement plus systématique des baies (Figure n° 2), même si des accidents de coulure (3*) peuvent encore arriver.


Diversification des formes et des adaptations
La sélection, depuis le début de la culture de la vigne jusqu’à aujourd’hui, a permis une diversification aussi bien sur des critères qualitatifs que sur des critères d’adaptation.
Ainsi, trois principaux types de variétés ont été créés : raisins de cuve, raisins de table, et raisins secs issus de schémas de sélection divergents. Pour les raisins secs, la sélection a porté sur l’apyrénie (4*) s’accompagnant de baies très petites, plus faciles à sécher. Pour les raisins de cuve et de table, elle a reposé principalement sur les baies et les grappes comme le montre l’étude des variétés de la collection du domaine de Vassal(5*) (Figure n° 3) : petites baies et petites grappes, maximisant le ratio surface de pellicule sur quantité de pulpe afin d’extraire un maximum de composés organoleptiques importants pour la qualité du vin pour la cuve ; grosses baies, grappe plus grande et caractères de diversification (arôme Muscat, apyrénie…) pour la table.
De même, la sélection réalisée par l’homme a permis d’adapter la vigne d’origine méditerranéenne à des zones de production variées allant, pour l’hémisphère nord, des vignobles allemands, anglais ou polonais les plus septentrionaux (> 51° N) aux vignobles marocains les plus au sud (< 28° N) et à certaines îles (Canaries, Santorin). Sous l’effet de la sélection, la phénologie de la vigne a ainsi évolué : dans la collection Inrae du Domaine de Vassal, on observe des variations sur la date de maturité de plus de quinze semaines, les variétés les plus précoces arrivant à maturité fin juillet, les plus tardives fin octobre (Figure n° 3).
En revanche, les variétés de Vitis vinifera utilisées aujourd’hui sont sensibles aux principales maladies de la vigne, à l’exception de certaines variétés résistantes au mildiou (par exemple portugaises), à l’oïdium (d’Asie centrale) ou au botrytis (cabernet sauvignon).

Ce que l’ADN nous a appris
De nouvelles informations ont été acquises grâce à l’analyse de l’ADN. Nous avons alors pu démontrer que, malgré une forte érosion génétique, il existe encore quelques populations de vignes sauvages. De même, l’analyse de la collection de Vassal a permis d’identifier trois pools génétiques : variétés de table du Moyen-Orient, variétés de cuve de l’Est et variétés de cuve de l’Ouest, démontrant l’effet de la sélection au niveau du génome. Nous avons également étudié la généalogie des cépages. Ainsi, le chardonnay et le gamay sont issus de croisements probablement spontanés entre le pinot noir et le gouais blanc. D’autres cépages, comme le savagnin et le gouais, sont les parents de nombreux descendants apparus sur une très longue période et désormais répartis dans le monde entier, démontrant l’existence d’importants échanges de matériel végétal depuis des temps reculés. Enfin, un gène impliqué dans l’avortement des organes femelles des fleurs a été récemment identifié et pourrait permettre d’expliquer le passage de la dioécie à l’hermaphrodisme chez la vigne. La vigne sauvage est une espèce protégée en France depuis 1995 et doit à tout prix être préservée. Elle pourrait être utilisée dans des programmes de création variétale visant l’amélioration de la vigne pour la qualité, la typicité et la tolérance au changement climatique.
Patrice This, Jean-Michel Boursiquot, Valérie Laucou, Jean-Pierre Péros et Thierry Lacombe
(1*) C’est-à-dire qu’il existe des pieds mâles et des pieds femelles.
(2*) Signifiant « vigne mariée aux arbres ».
(3*) Chute de fleur non ou mal fécondée.
(4*) Ou absence de pépins.
(5*) www6.montpellier.inrae.fr/vassal
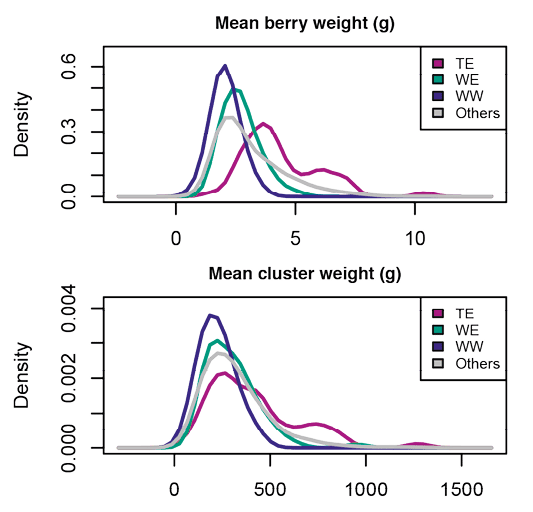
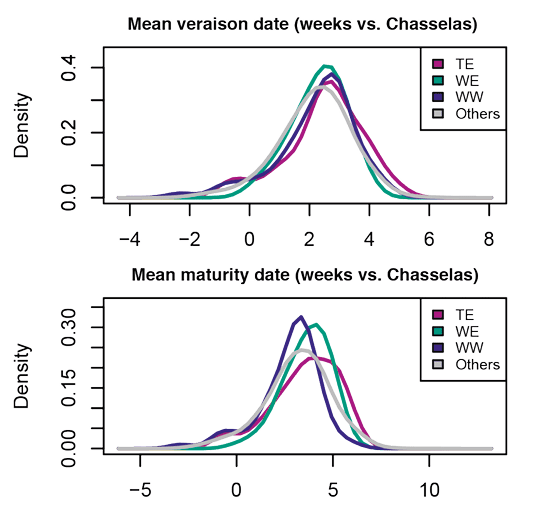
Figure n° 3: Distribution des poids de baie et de grappe (en grammes) et des dates moyennes de véraison et de maturité (en semaine par rapport à la variété Chasselas), pour la collection Inrae de Vassal et pour trois échantillons de vigne représentant les trois pools de la vigne: variétés de table du Moyen Orient (T. Est), variétés de cuve de l’Est (Cuve E) et variétés de cuve de l’Ouest (Cuve O). D’après Nicolas et al. 2016
À LIRE
Badouin H, Velt A, Gindraud F, Flutre T, Dumas V, Vautrin S, Marande W, Corbi J, Sallet E, Ganofsky J, Santoni S, Guyot D, Ricciardelli E, Jepsen J, Käfer J, Berges H, Duchêne E, Picard F, Hugueney P, Tavares R, Bacilieri R, Rustenholz C, Marais G. The Wild Grape Genome Sequence Provides Insights into the Transition from Dioecy to Hermaphroditism During Grape Domestication, Genome Biology. 2020;21:223
Bowers JE, Boursiquot JM, This P, Chu K, Johanson H, Meredith CP. Historical Genetics: the Parentage of Chardonnay, Gamay and other Wine Grapes of Northeastern France. Science. 199;285:1562-1565
Lacombe T, Boursiquot JM, Laucou V, Di Vecchi-Staraz M, Péros JP, This P. Large-Scale Parentage Analysis in an Extended Set of Grapevine Cultivars (Vitis vinifera L.). Theoretical Applied Genetics. 2013;126:401-414
Nicolas S, Peros JP, Lacombe T, Launay A, Le Paslier MC, Bérard A, Mangin B, Valière S, Martins F, Le Cunff L, Laucou V, Bacilieri
R, Dereeper A, Chatelet P., This P, Doligez A. Genetic Diversity, Linkage Disequilibrium and Power of a Large Grapevine (Vitis vinifera L) Diversity Panel Newly Designed for Association Studies. BMC Plant Biology 2016;16:74. DOI: 10.1186/s12870-016-0754-z Riaz S, Boursiquot JM, Dangl GS, Lacombe T, Laucou V, TenscherAC, Walker AM. Identification of Mildew Resistance
in Wild and Cultivated Central Asian Grape Germplasm.
BMC Plant Biology 2013;13:149. http://www.biomedcentral. com/1471-2229/13/149
This P, Bakry F, Froelicher Y, Luro F, Ollitrault P. Aspermie ou quand la graine n’est plus indispensable ou est indésirable ! « Les coulisses de la Floraison », Colloque Scientifique de la société Nationale d’Horticulture de France, Mai 2013, Rennes.
This P, Lacombe T, Thomas MR. Historical Origins and Genetic Diversity of Wine Grapes. Trends in Genetics 2006;22:511-519 This P, Roux C, Parra P, Siret R, Bourse T, Adam A-F, Yvon M, Lacombe T, David J et Boursiquot J-M. Caractérisation de la diversité d’une population de vignes sauvages du Pic Saint Loup (Hérault) et relations avec le compartiment cultivé. Genetics Selection Evolution. 2000;33:S289-S304
GLOSSAIRE
Nouaison : stade de développement de la baie correspondant à son développement précoce suite
à la fécondation de la fleur dont elle est issue.
Coulure : chute de fleur non ou mal fécondée, pouvant avoir différentes origines (physiologique, climatique, pathologique).
Véraison : changement physiologique de la baie de raisin correspondant au début du processus de maturation qui se traduit par son ramollissement et par son changement de couleur.